ニュース&イベント NEWS & EVENTS
新たな「細胞核のウイルス起源説」の提唱
~宿主ゲノムと"共存"して複製するという特徴をもった巨大ウイルスが、 細胞核の誕生のきっかけとなった可能性~
- ●真核生物の細胞核がどのように進化したかについては謎が多く、未だ定説がありません。
- ●本研究では、巨大ウイルス「メドゥーサウイルス」が、細胞核内で複製する唯一の巨大ウイルスであること、真核生物のヒストン遺伝子をフルセット持つことなど、真核生物との進化的な関わりを示唆する遺伝的特徴を有していることに加え、ここ十数年で明らかになってきた巨大ウイルスと真核生物の進化的関係に関する知見を総合し、真核生物の細胞核の進化を説明する新たな仮説を提唱しました。
- ●この新たな仮説は、細胞核の起源を説明する上で重要な2つの点、(1)ゲノムDNAの脂質二重層による包み込みがいかにして成り立ったのか、(2)リボソームはいかにして細胞核外へと排除されたのか、を明確に説明できており、細胞核の起源の解明に、一歩近づくと期待されます。
東京理科大学理学部第一部教養学科 武村政春教授は、2019年に日本の温泉水から分離した巨大ウイルス「メドゥーサウイルス」の遺伝的特徴や細胞核内で複製するその特性、ならびに巨大ウイルスの発見後に様々に議論されてきた細胞核の起源と、巨大ウイルスが宿主となるアメーバの細胞質内につくる「ウイルス工場」と呼ばれる特徴的な構造の進化的関係に関する知見を総合し、宿主のゲノムと"共存"して複製するという特徴をもったメドゥーサウイルスの祖先がきっかけとなり、宿主がウイルスに対する防御機構としてゲノムを囲う膜を構築したことが、細胞核の起源となったとする新たな「細胞核ウイルス起源説」を提唱しました。
本研究成果は、国際的なオンライン微生物学雑誌Frontiers in Microbiology誌に9月3日付けで掲載されました。
研究の背景
巨大ウイルスとは、いわゆる"ふつうの"ウイルスよりも粒子のサイズやゲノムサイズ、遺伝子数が大きく、「一部の機能を欠いた細胞性生物」とも思えるほど複雑なしくみをもったウイルスの総称で、「核細胞質性大型DNAウイルス(NCLDV)」と呼ばれる大型のDNAウイルス(注1)の仲間です。2003年に最初の巨大ウイルス「ミミウイルス」(注2)がヨーロッパで発見されて以来、数多くの巨大ウイルスが分離されてきました。日本からも、東京理科大学 武村教授を中心として、マルセイユウイルス科(注3)に含まれる「トーキョーウイルス」(東アジア・東南アジア地域ではじめて分離された巨大ウイルス)や、ミミウイルス科ウイルス、日本産パンドラウイルス、そしてどの系統にも属さないメドゥーサウイルスなどを水環境から分離してきました。一つの大きな特徴として、これらの巨大ウイルスのほとんどは、原生生物に感染します。
巨大ウイルスが注目される理由は、そのゲノムの起源の"古さ"にあります。巨大ウイルスの遺伝子を解析すると、その起源が真核生物(注4)の起源に近いところまで遡れることがわかっています。さらに、巨大ウイルスと真核生物の間では、これまで数多くの遺伝子水平移動(注5)が起こったとされ、とりわけメドゥーサウイルスと宿主であるアカントアメーバとの間には、そうした遺伝子が数多く存在することがわかっています。このように、巨大ウイルスの研究は、真核生物の進化の謎を紐解くカギとなると考えられています。
真核生物の進化の謎の中で、最も重要であるにも関わらず定説が存在しないのが、「細胞核の起源」です。これまでは、「細胞膜が内側に陥入し、核膜となった」という説明が高校生物の教科書でもなされていましたし、研究者もその考えに立脚して様々な細胞核起源説を提唱してきました。しかしこの説明では、(1)どのようなきっかけによってゲノムが核膜で包まれるようになったのか、そして(2)リボソームはなぜ細胞核の外側に排除されたのか、という細胞核の成立を説明するためになくてはならない2つの点を、満足に説明することはできてきませんでした。
2001年、当時まだ名古屋大学助手だった武村教授は、ポックスウイルスのような大型のDNAウイルスの感染が、細胞核の起源となったとする新たな仮説を提唱しました(注6)。その後、巨大ウイルスが発見され、巨大ウイルスのウイルス工場と細胞核との類似性が指摘され、さらに上述の、真核生物の進化の謎を解く鍵をもつと考えられるメドゥーサウイルスが発見されたことにより、このたび、従来の説では説明できなかった2つの点を明快に説明できる、細胞核の起源に関する新たな仮説を構築することに成功しました。
研究結果の概要
<メドゥーサウイルスの特徴に着目>
本研究で武村教授は、京都大学などとの共同研究により2019年に日本の温泉水から分離した「Acanthamoeba castellanii medusavirus(メドゥーサウイルス)」のいくつかの特徴に着目しました。一つは、ほかの巨大ウイルスとは異なり、メドゥーサウイルスのゲノムが宿主の細胞核全体に広がり、細胞核そのものをウイルス工場としているかのように複製する、ということです。このことは蛍光標識したメドゥーサウイルスDNAが宿主の細胞核全体で複製している様子を蛍光顕微鏡で捉えることに成功した先行研究(注7)において示されました。この研究ではDAPIという試薬でメドゥーサウイルス感染細胞のDNAを染色すると、細胞核が強く蛍光を発し、正常な細胞では見えていた核小体が消失し、まるでメドゥーサウイルスDNAにより細胞核が膨らんでいるようにも見えることがわかりました(図1a)。
一方、先行研究(注7)からメドゥーサウイルスの保有遺伝子の特徴も明らかになりました。メドゥーサウイルスにはほかの巨大ウイルスには存在する、遺伝子の転写や複製におけるDNAの立体構造維持にかかわる酵素など、いくつかの重要な遺伝子が存在せず、DNAポリメラーゼ、Ran、ヒストンなど、真核生物の遺伝子に非常によく似た遺伝子が存在することが明らかとなり、メドゥーサウイルスが細胞核で複製するに伴い、宿主の遺伝子に依存すれば事足りるものは失われ(Loss)、主要な遺伝子は何らかの理由で水平移動(lateral gene transfer:LGT)により宿主より獲得してきたと考えられました(図1b)。
これらのことから本研究では、メドゥーサウイルスの祖先となった巨大ウイルスが、宿主の細胞核において宿主ゲノムと"共存"して複製する戦略を編み出した結果、いくつかの遺伝子を失い、いくつかの遺伝子を水平移動により獲得したと考えました。現在のメドゥーサウイルスが、宿主との相同遺伝子を19%(この数字はほかの巨大ウイルスよりも格段に高い)も保有していることからも(注7の先行研究)、宿主ゲノムとの"共存"は、遺伝子の水平移動を推進してきたと考えられます。
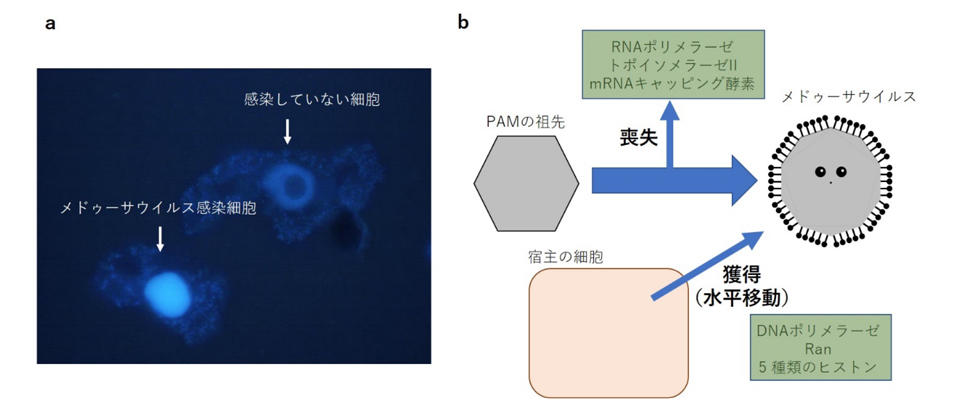 |
図1 メドゥーサウイルスの2つの特性。(a)メドゥーサウイルスは、宿主の細胞核全体を使って複製する。DNA染色試薬DAPIでDNAを染色し、蛍光顕微鏡で観察すると、メドゥーサウイルス感染細胞の細胞核が膨張したようにみえ、核小体がほぼ見えなくなることがわかった。(b)メドゥーサウイルスは、RNAポリメラーゼやトポイソメラーゼなど、ほかの巨大ウイルスには存在するいくつかの重要な遺伝子が存在せず、進化の過程で失ったと考えられる。
<細胞核の謎1・どのようなきっかけでゲノムが核膜で包まれるようになったのか>
それでは、そのメドゥーサウイルスと宿主のゲノム複製の場が、どのように脂質二重層で覆われるようになったのでしょうか。フランス・パスツール研究所のForterre博士らは、次のように考えています(注8)。(1)もともと細胞内部に進化していた小胞体や食胞などの細胞内膜系は、まずウイルスが利用し始め、宿主からのヌクレアーゼなどからの攻撃から身を守ろうとして、自らのゲノム複製の場(今でいうウイルス工場など)の周囲を囲わせるようになった。(2)宿主細胞も、そのウイルスの戦略を学んで、自らのゲノムの周囲を細胞内膜系で覆い、逆にウイルスからの攻撃から身を守ろうとするようになった、というのです。
バクテリアの細胞内に核膜のような膜がゲノムを囲うようにでき、それが細胞核へと進化した直接的な証拠はありませんが、感染したバクテリア内部に自らの複製工場をつくるバクテリオファージの存在や、細胞内部にゲノムを囲う内膜系を発達させているバクテリアの存在は、原核生物でもそのような内膜を形成し得ることを示唆しています。しかしながら、宿主ゲノムが内膜で包まれたことを説明するためには、ウイルスから身を守るためというdriving forceを考えただけでは、若干弱いと思われました。ただしそのウイルスが「メドゥーサウイルスの祖先」であったのなら、話は変わります。ゲノムと"共存"するように自らのDNAを複製するようなウイルスであったのなら、ゲノムを細胞内膜系で囲んでウイルス工場を作るというウイルス側の目的と、ゲノムを細胞内膜で囲んでゲノムを外敵から守るという細胞側の目的が、見事に合致するからです。
以上のことから武村教授は、細胞核の起源に関する重要な1つめの点、「どのようなきっかけによってゲノムが核膜で包まれるようになったのか」に関して、次のようなシナリオを構築しました。すなわち、宿主ゲノムと"共存"して複製する特徴をもった巨大ウイルス(おそらくメドゥーサウイルスの祖先)がきっかけとなり、そのウイルスに対する防御機構として宿主がゲノムを核膜で覆うという戦略を編み出した結果、巨大ウイルス側もそのまま、ウイルス工場として細胞核を用いる状態へと進化し、現在のメドゥーサウイルスが誕生したのです。
<細胞核の謎2・リボソームはなぜ細胞核の外側に排除されたのか>
それでは、2つめの点「リボソームはなぜ細胞核の外側に排除されたのか」は、どのように説明できるのでしょうか。
メドゥーサウイルスは、ほかの巨大ウイルスとは異なり、「Ran」と呼ばれるGTPase活性をもったタンパク質の遺伝子を保有しています。Ranタンパク質は、真核生物において、物質の細胞核と細胞質との間の輸送にかかわる重要なタンパク質で、核小体において組み立てられたリボソームサブユニットの細胞核内から細胞質への輸送にも関与しています。すなわち、メドゥーサウイルスの祖先が、核膜で覆われた自身のウイルス工場からリボソームを排除することが、宿主とウイルスの双方にとってメリットがあったため、リボソームの細胞核の外側への排除というしくみが出来上がったのです。
では、宿主とウイルスにとって、ウイルス工場からリボソームを排除するメリットとはどのようなものでしょうか。宿主もウイルスも、自らの遺伝子(のすべてあるいは一部)にイントロン(介在配列)が存在していることがわかっていますが、このイントロンを除去し、エキソン同士をつなげるスプライシングを行った後、別の場所に移動させ、そこでリボソームに翻訳させるという流れを作り出す必要があります(注9)。もしイントロンが除去されないままにリボソームで翻訳作業が行われると、欠陥品のタンパク質ができてしまうからです。したがって、核膜に覆われた内と外で、スプライシングと翻訳の場を別にすることに成功したことが、宿主とウイルス両方のメリットにつながったと考えられます(図2)。
以上のことから武村教授は、細胞核の起源に関する重要な2つめの点、「リボソームはなぜ細胞核の外側に排除されたのか」に関して、次のようなシナリオを構築しました。すなわち、イントロンが存在していなかった原核生物の時代には問題にならなかった問題が、イントロンが進化し、そのイントロンを有する宿主とウイルスの翻訳上の戦略において克服すべき問題となり、祖先型「Ran」を用いて、たまたまウイルス感染によって膜で囲まれる状態となっていたゲノムの領域の外へリボソームを排除するしくみを作り上げた結果、現在の真核生物の細胞核(メドゥーサウイルスのウイルス工場)が誕生したのです。
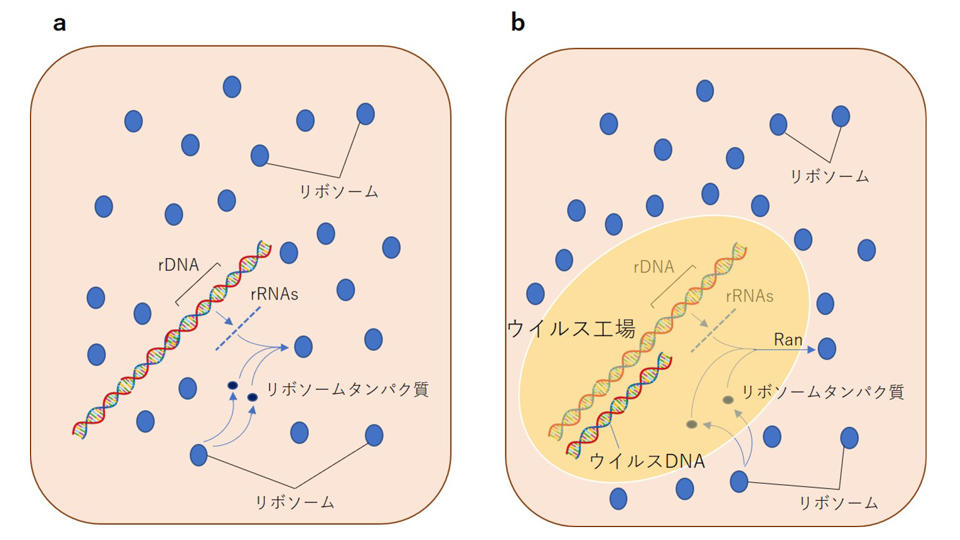 |
図2 リボソームの細胞核外への排除。左は細胞核成立前。右は成立後。ウイルス工場もしくは細胞核の成立とともに、Ranを用いたリボソーム排除のしくみが構築された。
<新たな細胞核のウイルス起源説>
以上のことから武村教授は、以下のシナリオにより細胞核が生まれたのではないかとする仮説を提唱しました。
(1) 現在存在する巨大ウイルスのうちPAMグループ(フィコドナウイルス科/アスファウイルス科/ミミウイルス科/モリウイルス・パンドラウイルス/メドゥーサウイルス)と呼ばれる巨大ウイルスの祖先が、真核生物の祖先(まだ細胞核はない)の細胞質に、細胞の小胞体に由来する内膜によって周囲を取り囲んだウイルス工場を形成していた(図3a)。
(2) 真核生物の祖先は、そのウイルス工場を模倣するかのように、ウイルスからの攻撃から身を守ろうとして、ゲノムの周囲を内膜で取り囲むしくみを進化させた。このゲノムを取り囲んだ内膜は一時的なものであるが、このころ、膜の中でゲノムを凝縮させるヒストンや、膜を通過させて物質輸送を行うRanなどの分子が進化した(図3b)。
(3) メドゥーサウイルスの祖先となったウイルスは、真核生物の祖先がゲノムを内膜で取り囲んだ状態へと進化しても、そのままゲノムと"共存"する形で複製をしていたため、そのために重要ないくつかの遺伝子、ヒストン、Ran、DNAポリメラーゼを水平移動により宿主から獲得した。その代わり、内膜で取り囲まれたことによってさらに宿主ゲノムへの依存を強くした結果、RNAポリメラーゼやトポイソメラーゼなどの重要遺伝子を欠失させた(図3b, c)。
(4) 真核生物の祖先において一時的にゲノムを取り囲んでいた内膜は、やがて恒久的にゲノムを取り囲む核膜へと進化し、メドゥーサウイルスの祖先となったウイルスにとってはそのまま、細胞核全体がウイルス工場として機能するようになった。同時に、Ranを使ってリボソームを外側へと排除するしくみが進化した(図3c)。
(5) 一方、ミミウイルスやマルセイユウイルスの祖先となったウイルスは、もともと宿主ゲノムとは離れたウイルス工場で複製していたため、核膜が進化してもその中に取り込まれず、そのまま細胞質のウイルス工場で複製する巨大ウイルスへと進化した。同様に、その他の巨大ウイルスもそれぞれに特有な複製メカニズムを進化させた(図3c)。
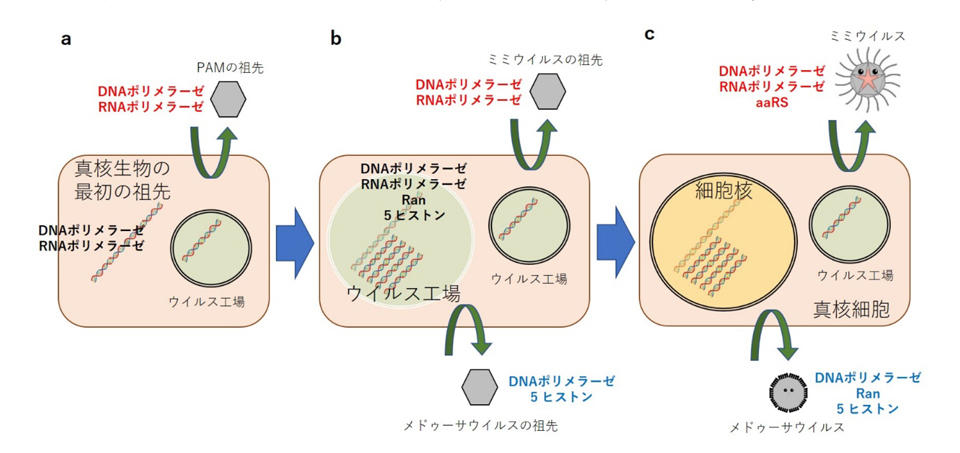 |
図3 新たな「細胞核のウイルス起源説」。黒字の遺伝子:宿主細胞の遺伝子。赤字の遺伝子:ミミウイルス科ウイルスの遺伝子。青字の遺伝子:メドゥーサウイルスの遺伝子。
今後の展望
細胞核がどのようにしてできたのかに関して、これまで多くの研究者が様々な仮説を提唱してきましたが、いずれも現象論にとどまります。代謝システムの進化という分子レベルの観点から細胞核の進化について言及しているものはありますが、より詳細にその「きっかけ」を考察したものはほとんどありません。小胞体を中心とした膜、もしくは共生したバクテリア(のちのミトコンドリア)の膜、などに由来する細胞内膜系が、複雑な過程を経て、やがてゲノムを包み込み、細胞核へと進化したというのは多くの研究者がそう考えていることですが、「やがてゲノムを包み込み」という一言が極めて重いものであるにもかかわらず、いずれも「ゲノムを包み込む」きっかけとなった重要なイベントについてはほとんど言及していませんでした。しかし、ウイルスの存在を仮定すると、本研究で述べてきたように、「ゲノムを包み込む」過程を非常に明快に、無理なく説明することができます。そして本研究により、細胞核の起源の解明に一歩近づくことが期待されます。
本研究では、メドゥーサウイルスのもつ特徴を織り交ぜた新しい細胞核の起源説を提唱しましたが、もちろんメドゥーサウイルスのみがカギを握るものではないでしょう。武村教授は今後、さらに様々なウイルスを分離し、そのデータをもとにさらに詳細な学説へと昇華させていきたいと考えています。
解説
注1 核細胞質性大型DNAウイルスとは、比較的大型のDNAウイルスで、ポックスウイルス科、フィコドナウイルス科、アスコウイルス科、アスファウイルス科、イリドウイルス科が含まれていたが、巨大ウイルスの発見により、ミミウイルス科、マルセイユウイルス科が追加された。基本的な特徴は、ほかのウイルスと同様にカプシドに覆われているが、その内側に脂質二重層があり、その中にゲノムDNAが存在することである。感染した細胞の細胞核もしくは細胞質で増殖するため、この名が付いている。
注2 ミミウイルスは、最初に発見された巨大ウイルスで、粒子径500 nm、ゲノムサイズ120万塩基対は、発見当時(2003年)最大であった。正式には「Acanthamoeba polyphaga mimivirus」である。それまでバクテリアの一種と思われていたため、「Mimicry virus」を略して「Mimivirus」と呼ばれる。
注3 マルセイユウイルス科は、ミミウイルス科以外ではじめて2009年に発見された巨大ウイルスの仲間である。正二十面体のカプシドをもつ"最小の巨大ウイルス"で、粒子径200 nm前後、ゲノムサイズは37万塩基対前後である。現在までにマルセイユウイルス、ローザンヌウイルス、メルボルンウイルス、チュニスウイルス、インセクトマイムウイルス、トーキョーウイルスなどが見つかっている。
注4 真核生物とは、私たちのように、細胞の中にゲノムDNAを含む細胞核を有する生物の総称。現在までに発見されている巨大ウイルスはすべて真核単細胞生物に感染する。
注5 遺伝子は、同種での親から子への移動(いわゆる遺伝のことで、垂直移動ともいう)以外に、別種同士の間で移動することがある。これを水平移動という。別種は交配することがないため、水平移動は基本的に生殖以外の方法で起こるが、そのうちの多くはウイルスを媒介した移動であると考えられている。そのためウイルスは"遺伝子の運び屋"とも呼ばれる。なお、ウイルスと宿主生物との間の水平移動も知られており、本研究で扱っているのはこのウイルス・宿主間の遺伝子水平移動である。
注6 ポックスウイルスの祖先が細胞核の誕生の元になったとする仮説で、TakemuraとBellにより2001年、国際分子進化雑誌『Journal of Molecular Evolution』誌上において独立に提唱され、Bellはこれを「Viral Eukaryogenesis」と名付けた。
注7 メドゥーサウイルスに関する以下の研究をさす。2019年に京都大学、ならびに東京理科大学からプレスリリースされ、科学誌『Nature』でも紹介された。
Yoshikawa G. et al (2019). Medusavirus, a novel large DNA virus discovered from hot spring water. J. Virol. 93, e02130-18, 2019.
注8 Forterre博士らの以下の研究をさす。
Forterre, P., and Prangishvili, D. (2009). The great billion-year war between ribosome- and capsid-encoding organisms (cells and viruses) as the major source of evolutionary novelties. Annu. N.Y. Acad. Sci. 1178, 65-77.
Forterre, P., and Gaïa, M. (2016). Giant viruses and the origin of modern eukaryotes. Curr. Opin. Microbiol. 31, 44-49.
注9 真核生物の遺伝子は、イントロンによって複数のエキソンに分断された状態になっているため、mRNAが転写された後、イントロン部分を除去する「スプライシング」と呼ばれる過程を経てから、リボソームで翻訳される必要がある。イントロンにはアミノ酸配列情報が存在しないため、除去されないまま翻訳されると、完全なタンパク質が合成されない。
論文情報
| 雑誌名 | : | Frontiers in Microbiology 2020年9月3日 オンライン掲載 |
|---|---|---|
| 論文タイトル | : | Medusavirus Ancestor in a Proto-eukaryotic Cell: Updating the Hypothesis for the Viral Origin of the Nucleus |
| 著者 | : | Masaharu Takemura |
| DOI | : | 10.3389/fmicb.2020.571831 |
武村研究室
研究室のページ:https://takemura-lab.azurewebsites.net/main/index
武村教授のページ:https://www.tus.ac.jp/fac_grad/p/index.php?4d94
東京理科大学について
東京理科大学:https://www.tus.ac.jp/
ABOUT:https://www.tus.ac.jp/info/index.html#houjin